
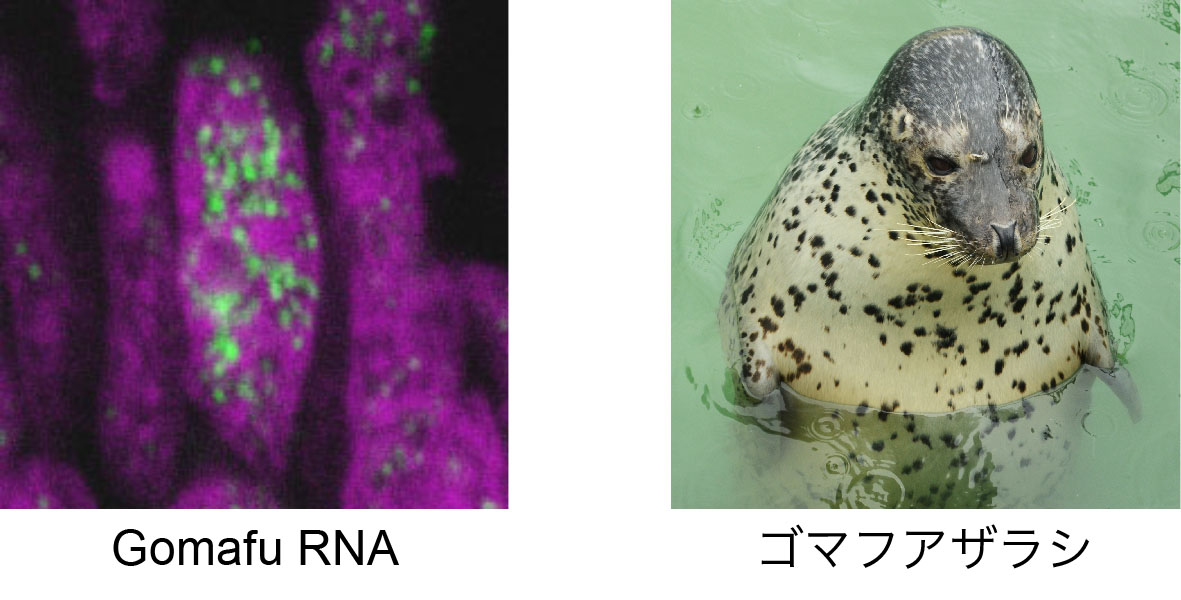
網膜の特定の細胞タイプで発現しているノンコーディングRNAとして同定されたGomafu、その細胞内局在がまるでゴマフアザラシなのでその名がついたGomafu。
 当時、発足直後の理研CDBで開始した僕のプロジェクトの目的は網膜の組織形成を制御する細胞表面の接着分子を同定することだったので、そのスクリーニングでたまたま取れてきたGomafuは核内の、しかもノンコーディングRNAということで、ラボ的にはランク外、ゴミ、カス、下手したら士気に対してはdominant negative扱い。とはいえ、なんで転写産物が他のmRNAのように細胞質に運ばれずに核にたまるのだろう、という素朴な疑問はずっと頭から離れず、まるで恋にとりつかれたように寝ても覚めても考えているのはGomafuのことばかり。同時並行で進めていた網膜の幹細胞の仕事や組織形成の仕事はそこそこ結果が出ていたもののそちらはすっかりおざなりになっているのを見かねた師匠からは、「私にはノンコーディングRNAの仕事がモノになるか分かりません」とやんわりと軌道修正を勧められていましたが、今はこんな扱いでもきちんとした生理機能が示せれば認めてもらえるはず、どんな頑固親父だって孫の顔を見りゃ、と猪突猛進でノックアウトマウス作りに突き進んでしまいました。普通、培養細胞を用いた実験である程度結果が出てから時間もコストもかかるノックアウトマウス作製のような実験に取り掛かるのがセオリーというものですが、神戸のCDBの変異マウスチーム(通称LARGE)というのがこれまた超スゴ腕で、当時マスターの学生さんだったHMさんでも10ラインぐらい平気で並行して変異マウスを作っていたので(それは彼女にド根性があったからなんでしょうが)、まあ、なんとかなるだろうと。
当時、発足直後の理研CDBで開始した僕のプロジェクトの目的は網膜の組織形成を制御する細胞表面の接着分子を同定することだったので、そのスクリーニングでたまたま取れてきたGomafuは核内の、しかもノンコーディングRNAということで、ラボ的にはランク外、ゴミ、カス、下手したら士気に対してはdominant negative扱い。とはいえ、なんで転写産物が他のmRNAのように細胞質に運ばれずに核にたまるのだろう、という素朴な疑問はずっと頭から離れず、まるで恋にとりつかれたように寝ても覚めても考えているのはGomafuのことばかり。同時並行で進めていた網膜の幹細胞の仕事や組織形成の仕事はそこそこ結果が出ていたもののそちらはすっかりおざなりになっているのを見かねた師匠からは、「私にはノンコーディングRNAの仕事がモノになるか分かりません」とやんわりと軌道修正を勧められていましたが、今はこんな扱いでもきちんとした生理機能が示せれば認めてもらえるはず、どんな頑固親父だって孫の顔を見りゃ、と猪突猛進でノックアウトマウス作りに突き進んでしまいました。普通、培養細胞を用いた実験である程度結果が出てから時間もコストもかかるノックアウトマウス作製のような実験に取り掛かるのがセオリーというものですが、神戸のCDBの変異マウスチーム(通称LARGE)というのがこれまた超スゴ腕で、当時マスターの学生さんだったHMさんでも10ラインぐらい平気で並行して変異マウスを作っていたので(それは彼女にド根性があったからなんでしょうが)、まあ、なんとかなるだろうと。
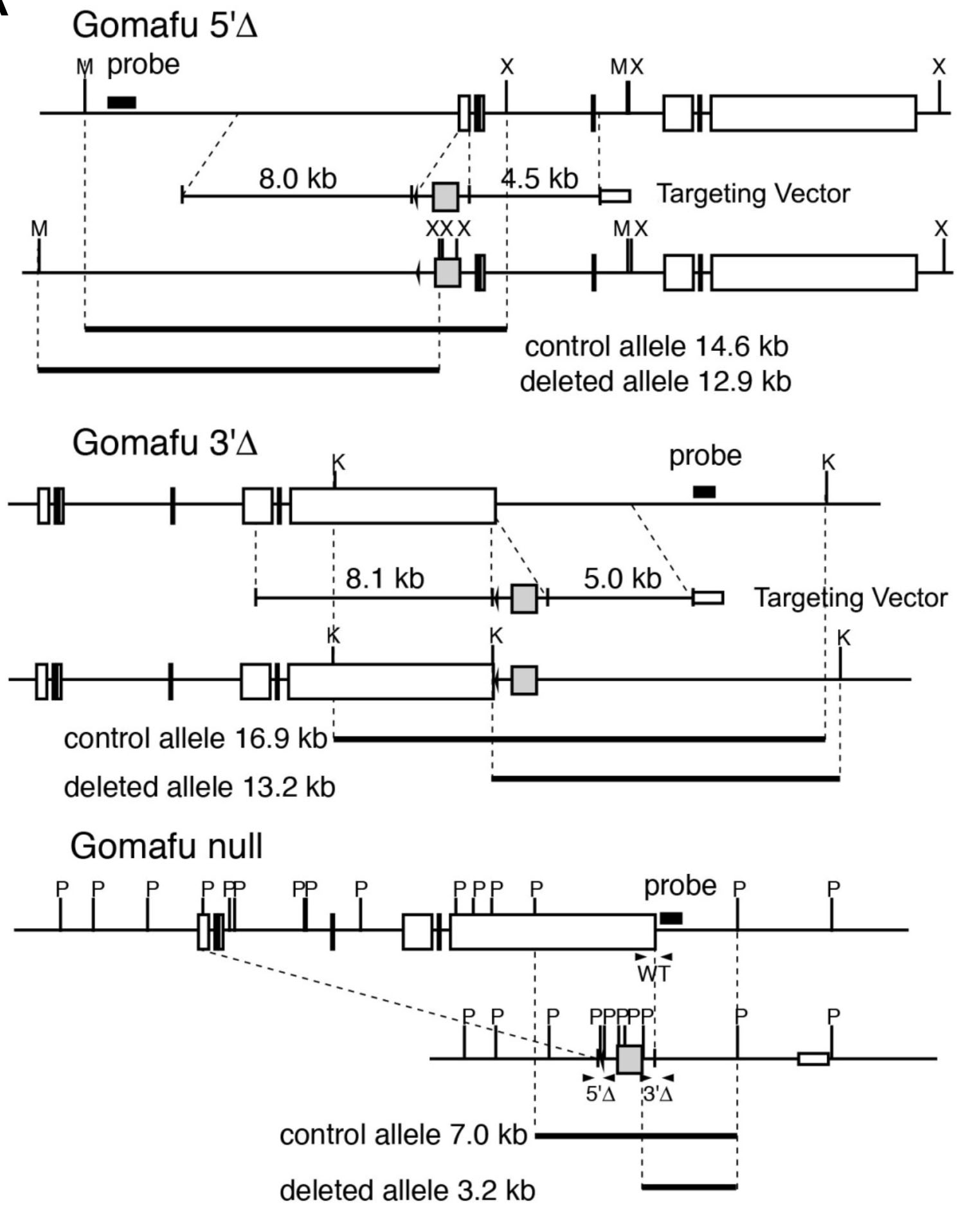
今であればCRISPR-Cas9を用いて遺伝子座の両端をチョクチョキすればすぐnull変異体が作れるのでしょうが、相同組換えを用いたやり方ではせいぜい削れるのはLARGEスタッフをもってしても5 kbぐらいとのことでした。GomafuはmRNAでも9 kb、遺伝子となると15 kbもあります。エクソンの大部分を削ったとしても、機能が残っていたら表現型は出なくなってしまいます。どうしようかと思い悩んでいたところ、Hox遺伝子座を自遊自在に削って様々な解析をしているDenis Dubouleさんのセミナーで、Cre-loxPのシステムを利用したTAMEREという手法があることを知りました。通常、別々の相同染色体上にあるloxP配列の間ではCre組換え酵素の存在下でも距離的に遠すぎて組換えは起きないのですが、減数分裂期には相同染色体が至近距離に接近するので、ある確率であたかも同じ染色体に乗っているloxP配列のように組換えが起きる、というとっても賢い方法です。このやり方を使えば、Gomafuの5'側と3'側にそれぞれloxP配列をノックインしたマウスを2ライン作り、それらをトランスヘテロに持つ個体の精原細胞でCreを発現させてやれば、スコンとGomafuの全領域を抜いてやることができます。
 ちなみに、このTAMEREなる方法のミソとなる精原細胞でCreを発現するラインを開発したのは僕がLong noncoding RNAのKeystoneのミーティングで朝食をとっている時にコップの水が気道に入ってゲホゲホ!ゲボボボッツッグッッウと死にそうになってしまった時にたまたま隣に居合わせて背中をトントンしていただいた優しい
ちなみに、このTAMEREなる方法のミソとなる精原細胞でCreを発現するラインを開発したのは僕がLong noncoding RNAのKeystoneのミーティングで朝食をとっている時にコップの水が気道に入ってゲホゲホ!ゲボボボッツッグッッウと死にそうになってしまった時にたまたま隣に居合わせて背中をトントンしていただいた優しいおばちゃまお姉さま箕面さん、でなくてMinoo Rassoulzadeganさんと知ったのはつい最近のことでありました。世の中狭し。。。
その後、2005年に和光でラボを立ち上げる機会をいただき、これからは誰にも遠慮なくncRNAの仕事ができる!!!と意気揚々とGomafuの仕事にどっぷりと浸かり、最初のノックアウトマウスのホモ個体が生まれてきたのはラボ発足直後2006のことでした。
生まれてきた、、、
そう。この時から。lncRNAのノックアウトマウスを作れども作れども外見ピンピンしているという、賽の河原の石積みが始まったのはちょうど10年前のことになります。(つづく)

 2020
2020